Регулирање на синтезата на холестерол
Регулацијата на синтезата на холестерол - неговиот клучен ензим (HMG-CoA редуктаза) се спроведува на различни начини.
Фосфорилација / дефосфорилација на HMG редуктаза. Со зголемување на односот на инсулин / глукагон, овој ензим се фосфорилира и преминува во активна состојба. Дејството на инсулин се спроведува преку 2 ензими.
HMG-CoA редуктаза киназа фосфатаза, која ја претвора киназата во неактивна дефосфорилизирана состојба:
Фосфотаза HMG-Co редуктаза со претворање во дефосфорилизирана активна состојба. Резултатот од овие реакции е формирање на дефосфорилизирана активна форма на редуктаза HMG-CoA.
Како резултат на тоа, за време на периодот на апсорпција, холестеролот се зголемува. Во овој период се зголемува достапноста на почетната подлога за синтеза на холестерол - ацетил - CoA (како резултат на јадење храна која содржи јаглехидрати и масти, бидејќи CoA ацетилот се формира при распаѓање на гликоза и масни киселини).
Во постсасорбитната состојба, глукагонот преку протеиногеназа А ја стимулира фосфорилацијата на HMG - CoA - редуктаза, претворајќи го во неактивна состојба. Оваа акција е засилена со фактот дека во исто време глукагонот ја стимулира фосфорилацијата и инактивацијата на фосфотазата на HMG-CoA редуктазата, со што ја задржува HMG-CoA редуктазата во фосфорилизирана неактивна состојба. Како резултат, синтезата на холестерол во периодот по апсорпција и за време на постот е инхибирана со ендогена синтеза. Ако содржината на холестерол во храната беше доведена до 2%, тогаш нагло се намали синтезата на ендоген холестерол. Но, целосното престанување на синтезата на холестерол не се јавува.
Степенот на инхибиција на биосинтезата на холестерол под влијание на холестерол што доаѓа од храна варира од личност до личност. Ова укажува на индивидуалноста на процесите на формирање на холестерол. Со намалување на интензитетот на синтезата на холестерол, можно е да се намали неговата концентрација во крвта.
Ако се расипе рамнотежата помеѓу внесот на холестерол со храна и неговата синтеза во организмот од една страна и излачувањето на жолчните киселини и холестеролот од друга страна, се менува концентрацијата на холестерол во ткивата и крвта. Најсериозните последици се поврзани со зголемување на концентрацијата на холестерол во крвта (хиперхолестеролемија), додека веројатноста за развој на атеросклероза и холелитијаза се зголемува.
Семејната хиперхолестеролемија (HCS) - оваа форма е многу почеста - околу 1 пациент на 200 луѓе. Наследниот дефект на HCS е повреда на апсорпцијата на LDL од страна на клетките, а со тоа и намалување на стапката на LDL катаболизам. Како резултат на тоа, се зголемува концентрацијата на ЛДЛ во крвта, како и холестерол, бидејќи има многу од тоа во ЛДЛ. Затоа, со HCS, карактеристично е таложење на холестерол во ткивата, особено во кожата (ксантоми), во theидовите на артериите.
Инхибиција на синтезата на HMG-CoA редуктаза
Конечен производ на метаболички пат на холестерол. Ја намалува стапката на транскрипција на генот HMG-CoA редуктаза, со што се инхибира сопствената синтеза. Црниот дроб активно синтетизира жолчни киселини од холестерол, и затоа жолчните киселини ја инхибираат активноста на генот HMG-CoA редуктаза. Бидејќи HMG-CoA редуктазата постои по синтеза на околу 3, инхибицијата на синтезата на овој ензим на холестерол е ефикасна регулатива.
Не го најдовте она што го баравте? Користете го пребарувањето:
Размена на холестерол естер
Фондот за холестерол содржи бесплатни хостери на холестерол и холестерол, кои се наоѓаат како во клетките, така и во липопротеините во крвта.
Дел II Метаболизам и енергија
Во клетките, естерификацијата на холестеролот се јавува со дејство на ацил-КоА-холестерол-ацилтрансфераза (АХАТ):
Ацил-CoA + холестерол - * HS-KoA + ацилхолестерол
Во човечките клетки, главно се формира линолејхолестерол. За разлика од бесплатниот холестерол, неговите естри во клеточните мембрани се содржани во многу мали количини и главно се наоѓаат во цитосолот како дел од капките на липидите. Формирањето на естри може да се смета, од една страна, како механизам за отстранување на вишокот холестерол од мембраните, а од друга страна, како механизам за чување холестерол во клетката. Мобилизацијата на ресурсите се јавува со учество на ензими на естераза кои ги хидролизираат естерите на холестерол:
Ацилхолестерол + Н 2О - * масна киселина + холестерол
Синтезата и хидролизата на естерите се јавува кај многу клетки, но е особено активна кај клетките на надбубрежниот кортекс: кај овие клетки до 80% од целиот холестерол е претставена со естри, додека во други клетки обично е помала од 20%.
Формирањето на естри во липопротеините во крвта се одвива со учество на лецитин-холестерол ацилтрансфераза (LHAT), што го катализира трансферот на ацилскиот остаток од i-позицијата на лецитин во холестерол (Сл. 10.31). LHAT се формира во црниот дроб, се излачува во крвта и се прикачува на липопротеините. Стапката на естерификација за различни липопротеини е значително различна и зависи од присуството на аполипротеини кои го активираат LHAT (главно апо-АТ, како и CI) или го инхибираат (C-II) овој ензим.Најктивниот LHAT во HDL, во кој апо-АТ мора да olee 2/3 од сите протеини. На најголемиот coli почести формирана естри на олеинска и линолова киселини. На други липопротеини естер на формирање се случува со помала стапка отколку во HDL.
Сл. 10.31. Формирање на холестерол естери под дејство на LHAT
LHAT е локализиран во површинскиот слој на HDL и како супстрат користи холестерол во фосфолипидниот монослоен. Хостемите на холестерол формирани тука, заради нивната целосна хидрофобност, слабо се задржуваат
Поглавје 10. Метаболизам и липидна функција
фосфолипиден монослоен и потопен во липидното јадро на липопротеинот. Во исто време, во фосфолипидниот монослоен се ослободува место за холестерол, кое може да се наполни со холестерол од клеточните мембрани или од други липопротеини. Така, HDL се чини дека е стапица на холестерол како резултат на дејството на LHAT.
Синтеза на жолчката киселина
Во црниот дроб, дел од холестеролот се претвора во жолчни киселини. Галните киселини можат да се сметаат за деривати на холанска киселина (Сл. 10.32).
Холанска киселина како таква не е формирана во организмот. Кај хепатоцитите, холестеролот директно произведува фенодеоксихолни и холични киселини - примарни жолчни киселини (Сл. 10.33, видете исто така на Сл. 10.12).
Биосинтеза на холестерол
Биосинтезата на холестерол се јавува во ендоплазматскиот ретикулум. Изворот на сите атоми на јаглерод во молекулот е ацетил-СКОА, кој доаѓа од митохондриите како дел од цитрат, како во синтезата на масни киселини. Биосинтезата на холестерол троши 18 АТП молекули и 13 молекули на NADPH.
Формирањето на холестерол се јавува во повеќе од 30 реакции, кои можат да се групираат во неколку фази.
1. Синтеза на мевалонска киселина.
Првите две реакции на синтеза се совпаѓаат со реакциите на кетогенеза, но по синтезата на 3-хидрокси-3-метилгутарил-ScoA, влегува ензимот хидроксиметил-глутарил-ScoA редуктаза (HMG-SCOA редуктаза), формирајќи мевалонска киселина.
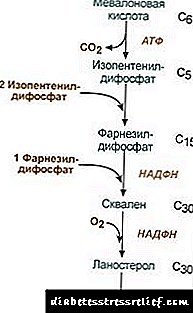 |
Шема на реакција на синтеза на холестерол2. Синтеза на изопентенил дифосфат. Во оваа фаза, три остатоци од фосфат се прикачени на мевалонска киселина, потоа е депарбоксилизирана и дехидрогенирана. 3. По комбинирање на трите молекули на изопентенил дифосфат, се синтетизира фарнезил дифосфат. 4. Синтезата на сквален се јавува кога се врзани два остатоци на фарнезил дифосфат. 5. По сложени реакции, линеарен сквален циклира до ланостерол. 6. Отстранување на вишок метилни групи, реставрација и изомеризација на молекулот доведува до појава на холестерол. Регулирање на активноста на хидроксиметилглутарил-С-CoA редуктаза3. Стапката на биосинтеза на холестерол исто така зависи од концентрацијата специфичен протеински носачобезбедување на врзување и транспорт на метаболити со хидрофобна средно синтеза. Може да го прашате или оставите вашето мислење. Главната точка на регулирање е реакцијата на формирање на мевалонска киселина. 1. Алостерско регулирање. Холестерол, и во црниот дроб - и жолчните киселини ја инхибираат редуктазата на HMG-CoA. 2. Репресија на синтезата на HMG-CoA редуктаза холестерол. 3. Регулација со фосфорилација-дефосфорилација на HMG-CoA редуктаза, активна нефосфорилизирана форма. Глукагонот предизвикува деактивирање, а инсулин предизвикува активирање преку сложена каскада на реакции. Така, стапката на синтеза на холестерол се менува со промена на состојбите со апсорпција и постсорпција. 4. Стапката на синтеза на HMG-CoA редуктаза во црниот дроб е предмет на јасни дневни флуктуации: максимум на полноќ, а минимум наутро. Размена на холестерол естер Во клетките естерификација на холестерол се јавува кога се изложени ацил-CoA-холестерол-ацилтрансфераза (АХАТ): Ацил-CoA + холестерол ® NS-CoA + Ацилхолестерол Во клетките, главно, се формира линолејхолестерол. Естерите се наоѓаат главно во цитосолот како дел од липидните капки. Формирањето на естри може да се смета, од една страна, како механизам за отстранување на вишокот холестерол од мембраните, а од друга страна, како механизам за чување холестерол во клетката. Мобилизацијата на резервите се одвива со учество на ензими естеразахидролизирање на естери на холестерол: Ацилхолестерол + Н2О ® масна киселина + холестерол Синтезата и хидролизата на естерите е особено активна во клетките на надбубрежниот кортекс. Кај липопротеините во крвта формирање на естер се јавува со учество на лецитин-холестерол-ацилтрансфераза (LHAT), катализирајќи го трансферот на остатоци од ацил од лецитин во холестерол. LHAT се формира во црниот дроб, се лачи во крвотокот и е прикачен на липопротеини. Најактивниот LHAT во HDL, каде што е локализиран во површинскиот слој. Хостерите на холестерол формирани тука се хидрофобни и потопени во липидното јадро. Во фосфолипидниот монослоен, има слободен простор за холестерол, кој може да се наполни со холестерол од клеточните мембрани или од други липопротеини. Така, HDL се чини дека е стапица на холестерол како резултат на дејството на LHAT. Синтеза на жолчката киселина Во црниот дроб, дел од холестеролот се претвора во жолчни киселини. Bолчните киселини може да се сметаат за деривати на холанска киселина. Холанска киселина како таква не е формирана во организмот. Кај хепатоцитите од холестерол се формираат примарни жолчни киселини - фенодексихоличен и холичен. После секрецијата на жолчката во цревата под дејство на ензими на цревната флора, од нив се формираат секундарни жолчни киселини - литохолски и деоксихоличен. Тие се апсорбираат од цревата, со крвта на порталната вена влегуваат во црниот дроб, а потоа и во жолчката. Bолчката содржи главно конјугирани жолчни киселини, т.е., нивните соединенија со глицин или таурин. Концентрацијата на жолчните киселини во жолчката е околу 1%. Вклучен е главниот дел на жолчните киселини хепатоцентрична циркулација.Мал дел од жолчните киселини - околу 0,5 g на ден - се излачува во измет. Ова се компензира со синтезата на нови жолчни киселини во црниот дроб, фондот на жолчна киселина се ажурира за околу 10 дена. Холестеролот исто така се излачува главно преку цревата. Влегува во цревата со храна и од црниот дроб како дел од жолчката. Холестеролот што се апсорбира во крвта содржи дел што потекнува од жолчката (ендоген холестеролсинтетизираат во црниот дроб) и фракцијата добиена од храна (егзогени холестерол) Отстранувањето на холестерол од ткива се јавува со негова оксидација на жолчните киселини во црниот дроб, проследено со нивно излачување со измет (приближно 0,5 g на ден) и со излачување на непроменет холестерол (исто така со измет). Во стационарна состојба: (Холестеролкрај + Холестеролпр) - (холестеролизмет + Bолчни киселиниизмет) = 0 Ако оваа рамнотежа е нарушена, концентрацијата на холестерол во ткивата и во крвта се менува. Зголемен холестерол во крвта - хиперхолестеролемија. Ова ја зголемува веројатноста за појава на атеросклероза и жолчни камења. РЕГУЛАТУВА LЕ НА ЛИПИДНИТЕ РАЗВОЈ Метаболизмот на липидите е регулиран од страна на централниот нервен систем. Долготрајна негативен емоционален стрес, зголемувањето на ослободување на катехоламини во крвотокот може да предизвика забележително слабеење. Акција глукагон на липолитичкиот систем е слично на дејството на катехоламини. Адреналин и норепинефрин ја зголемуваат активноста на липазата на ткивата и стапката на липолиза во масното ткиво, како резултат, се зголемува содржината на масни киселини во крвната плазма. Инсулин има спротивен ефект на адреналин и глукагон врз липолизата и мобилизацијата на масните киселини. Хормон за раст ја стимулира липолизата, поттикнувајќи ја синтезата на аценилат циклаза. Хипофизата хипофизата доведува до таложење на маснотии во телото (дебелина на хипофизата). Тироксин, полови хормонивлијаат и врз липидниот метаболизам. Отстранувањето на половите жлезди кај животните предизвикува вишок таложење на маснотии. РЕПУБЛИКА РАЗМЕНА НА ЛИПИД Холестеролот е стероид специфичен за животинските организми. Главното место на неговото формирање во човечкото тело е црниот дроб, каде што се синтетизира 50% холестерол, 15-20% се формира во тенкото црево, а остатокот се синтетизира во кожата, надбубрежниот кортекс и гонадите. Изворите на формирање на холестерол фонд и начините на неговото трошење се прикажани на Сл. 22.1. Сл. 22.1. Формирање и дистрибуција на холестерол во организмот. Холестеролот на човечкото тело (вкупна количина од околу 140 g) условно може да се подели на три базени: 30 g), што брзо се разменува, се состои од холестерол на цревниот ид, крвна плазма, црн дроб и други паренхимски органи, обновувањето се јавува за 30 дена (1 g / ден), 50 g), полека разменувајќи холестерол на други органи и ткива, 60 g), многу бавно разменуван холестерол на 'рбетниот мозок и мозокот, сврзно ткиво, стапката на ажурирање се пресметува со годините. Синтезата на холестерол се јавува во цитозолот на клетките. Ова е една од најдолгите метаболички патеки во човечкото тело. Продолжува во 3 фази: првиот завршува со формирање на мевалонска киселина, втората со формирање на сквален (линеарна структура на јаглеводород се состои од 30 јаглеродни атоми). Во текот на третата фаза, скваленот се претвора во молекула ланостерол, тогаш има 20 последователни реакции кои го претвораат ланостеролот во холестерол. Во некои ткива, хидроксилната група на холестерол естерифицира за да формира естри. Реакцијата се катализира со интрацелуларен ензим AHAT (ацилКоА: холестерол ацилтрансфераза). Реакцијата на естерификација се јавува и во крвта во ХДЛ, каде се наоѓа ензимот LHAT (лецитин: холестерол ацилтрансфераза). Хостемите на холестерол се форма во која се транспортираат со крв или се депонираат во клетките. Во крвта, околу 75% од холестеролот е во форма на естри. Синтезата на холестерол се регулира со влијание на активноста и количината на клучниот ензим на процесот - 3-хидрокси-3-метилгултарил-CoA редуктаза (HMG-CoA редуктаза). Ова се постигнува на два начина: 1. Фосфорилација / дефосфорилација на редуктаза HMG-CoA. Инсулинот ја стимулира дефосфорилацијата на HMG-CoA редуктазата, а со тоа преведувајќи ја во активна состојба. Затоа, во периодот на апсорпција, синтезата на холестерол се зголемува. Во овој период, исто така се зголемува достапноста на почетната подлога за синтеза, ацетил-CoA. Глукагонот има спротивен ефект: преку протеинска киназа А, ја стимулира фосфорилацијата на HMG-CoA редуктазата, претворајќи го во неактивна состојба. Како резултат, синтезата на холестерол во периодот по апсорпција и за време на постот е инхибирана. 2. Инхибиција на синтезата на HMG-CoA редуктаза.Холестеролот (краен производ на метаболичката патека) ја намалува брзината на транскрипција на генот HMG-CoA редуктаза, со што ја инхибира сопствената синтеза, а жолчните киселини исто така предизвикуваат сличен ефект. Транспортот на холестерол во крвта се изведува како дел од лекот. ЛП обезбедуваат егзогени холестерол во ткивата, ги одредуваат неговите протоци помеѓу органите и излачувањето од телото. Егзогениот холестерол се доставува до црниот дроб како дел од преостаната CHM. Таму, заедно со синтетизираниот ендоген холестерол, формира заеднички фонд. Кај хепатоцитите, ТАГите и холестеролот се пакуваат во VLDL, а во оваа форма се лачат во крвта. Под влијание на ЛП-липаза, хидролизирајќи го ТАГ до глицерол и масни киселини во крвта, VLDLPs прво се претвораат во СТЛП, а потоа во ЛДЛП кои содржат до 55% холестерол и негови естри. ЛДЛ е главната транспортна форма на холестерол во која се доставува до ткива (70% холестерол и неговите естри во крвта се дел од ЛДЛ). LDL од крвта влегува во црниот дроб (до 75%) и други ткива кои имаат LDL рецептори на нивната површина. Ако количината на холестерол што влегува во клетката ја надминува нејзината потреба, тогаш е задушена синтезата на LDL рецепторите, со што се намалува протокот на холестерол од крвта. Со намалување на концентрацијата на слободен холестерол во клетката, напротив, се активира синтеза на рецептори. Хормоните се вклучени во регулирањето на синтезата на LDL рецепторот: инсулин, тријодотиронин и половите хормони го зголемуваат формирањето на рецептори, а глукокортикоидите се намалуваат. Во таканаречениот „транспорт за враќање на холестеролот“, т.е. патеката што обезбедува враќање на холестеролот во црниот дроб, ХДЛ ја игра главната улога. Тие се синтетизираат во црниот дроб во форма на незрели претходници кои практично не содржат холестерол и ТАГ. HD прекурсорите во крвта се заситени со холестерол, примајќи го од други ЛП и клеточни мембрани. Пренесувањето на холестерол во ХДЛ вклучува ензим LHAT лоциран на нивната површина. Овој ензим прикачува остаток на масна киселина од фосфатидилхолин (лецитин) до холестерол. Како резултат, се формира хидрофобичен молекул на холестерол естер, кој се движи внатре во HDL. Така, не пијан HDL, збогатен со холестерол, се претвора во HDL 3 - зрели и поголеми честички. HDL 3 разменувајте естери на холестерол за TAG содржани во VLDL и СПБ со учество на специфичен протеин кој ги пренесува холестерол естерите помеѓу липопротеините. Во овој случај, HDL 3 се претвори во HDL2, чија големина се зголемува како резултат на акумулацијата на TAG. VLDL и HDLP под влијание на ЛП-липаза се претвораат во ЛДЛ, кои главно даваат холестерол на црниот дроб. Мал дел од холестерол се доставува до црниот дроб на HDL2 и HDL. Синтеза на жолчни киселини. Во црниот дроб, 500-700 мг жолчни киселини на ден се синтетизираат од холестерол. Нивното формирање вклучува реакции при воведувањето хидроксилни групи со учество на хидроксилази и реакција на делумна оксидација на страничниот ланец на холестерол (Сл. 22.2): Сл. 22.2. Шема на формирање на жолчни киселини. Првата реакција на синтеза - формирање на 7-а-хидроксихолестерол - е регулаторна. Активноста на ензимот што ја катализира оваа реакција е инхибирана од крајниот производ на патеката, жолчните киселини. Друг механизам за регулирање е фосфорилација / дефосфорилација на ензимот (активна е фосфорилираната форма на 7-а-хидроксилаза). Регулацијата е исто така можна со промена на количината на ензимот: холестеролот предизвикува транскрипција на генот 7-а-хидроксилаза и репресираат жолчните киселини. Тироидните хормони предизвикуваат синтеза на 7-а-хидроксилаза и репресија на естроген. Ваквиот ефект на естрогенот врз синтезата на жолчните киселини објаснува зошто болеста на жолчни камења се јавува кај жени 3-4 пати почесто отколку кај мажите. Холичните и фенодексихоличните киселини формирани од холестерол се нарекуваат „примарни жолчни киселини“. Најголемиот дел од овие киселини се подложени на конјугација - додавање на молекули на глицин или таурин во карбоксилната група на жолчна киселина. Конјугацијата започнува со формирање на активна форма на жолчни киселини - деривати на CoA, потоа се прикачени таурин или глицин, а како резултат се формираат 4 варијанти на конјугати: таурохолски и таурохенодеоксихолични, гликохолни и гликоенодексихолични киселини. Тие се значително посилни емулгатори од оригиналните жолчни киселини. Конјугатите со глицин се формираат 3 пати повеќе отколку со тауринот, бидејќи количината на таурин во организмот е ограничена. Во цревата, мала количина на конјугати на примарни жолчни киселини под дејство на бактериски ензими се претвораат во секундарни жолчни киселини. Деоксихолна киселина, формирана од холична и литохолна киселина, формирана од деоксихолна, е помалку растворлива и полека се апсорбира во цревата. Околу 95% од жолчните киселини кои влегуваат во цревата се враќаат во црниот дроб преку порталната вена, потоа повторно се излачуваат во жолчката и повторно се употребуваат во емулгацијата на мастите. Овој пат на жолчните киселини се нарекува ентерохепатична циркулација. Со измет, секундарните жолчни киселини главно се отстрануваат. Болеста на жолчното кесе (холелитијаза) е патолошки процес во кој се формираат камења во жолчното кесе, чија основа е холестерол. Ослободувањето на холестерол во жолчката треба да биде придружено со пропорционално ослободување на жолчни киселини и фосфолипиди, кои ги одржуваат молекулите на хидрофобен холестерол во мицеларна состојба. Причините што доведуваат до промена во односот на жолчните киселини и холестеролот во жолчката се: храна богата со холестерол, висококалорична исхрана, стагнација на жолчката во жолчното кесе, нарушена ентерохепатална циркулација, нарушена синтеза на жолчни киселини, инфекција на жолчното кесе. Кај повеќето пациенти со холелитијаза се зголемува синтезата на холестерол, а синтезата на жолчните киселини од него е забавена, што доведува до диспропорција во количината на холестерол и жолчни киселини излачувани во жолчката. Како резултат, холестеролот започнува да се таложи во жолчното кесе, формирајќи вискозен талог кој постепено се стврднува. Понекогаш тоа е импрегнирано со билирубин, протеини и соли на калциум. Камените можат да се состојат само од холестерол (камења на холестерол) или мешавина од холестерол, билирубин, протеини и калциум. Холестеролските камења обично се бели, а мешаните камења се кафеави во различни нијанси. Во почетната фаза на формирање на камен, фенодеоксихолна киселина може да се користи како лек. Откако ќе се најде во жолчното кесе, постепено ги раствора холестеролските камења, но ова е бавен процес кој трае неколку месеци. Атеросклерозата е патологија која се карактеризира со појава на атерогени плаки на внатрешната површина на васкуларниот ид. Една од главните причини за развој на таква патологија е нарушување на рамнотежата помеѓу внесот на холестерол од храна, неговата синтеза и екскреција од организмот. Пациентите со атеросклероза имаат зголемени концентрации на LDL и VLDL. Постои инверзна врска помеѓу концентрацијата на ХДЛ и веројатноста за развој на атеросклероза. Ова е во согласност со концептот на функционирање на ЛДЛ како носители на холестерол во ткивата и ХДЛ од ткива. Основен метаболички „предуслов“ за развој на атеросклероза е хиперхолестеролемија. (висок холестерол во крвта). Се развива хиперхолестеролемија: 1. поради вишок внес на холестерол, јаглехидрати и масти, 2. генетска предиспозиција која се состои во наследна дефект во структурата на LDL или апоБ-100 рецептори, како и во зголемена синтеза или секреција на apoB-100 (во случај на фамилијарна комбинирана хиперлипидемија, во која се покачени концентрациите на крвта и холестеролот и ТАГ). Важна улога во механизмите на развој на атеросклероза се игра со модификација на лекот. Промените во нормалната структура на липидите и протеините во ЛДЛ ги прават странски за организмот и затоа се подостапни за фаќање од фагоцити. Модификација на лекови може да се случи со неколку механизми: 1. гликозилација на протеини што се јавува кога се зголемува концентрацијата на глукоза во крвта, 2. модификација на пероксид, што доведува до промени во липидите во липопротеините и структурата на апоБ-100, 3. формирање на автоимуни комплекси на ЛП-антитело (променети лекови може да предизвикаат формирање на автоантитела). Изменетата ЛДЛ се апсорбира од макрофаги. Овој процес не е регулиран со количината на апсорбирана холестерол, како во случај на нејзино влегување во клетки преку специфични рецептори, затоа макрофагите се преоптоварени со холестерол и се претвораат во „пенести клетки“ кои продираат во субентелијалниот простор. Ова доведува до формирање на липидни дамки или ленти во theидот на крвните садови. Во оваа фаза, васкуларниот ендотелум може да ја одржи својата структура. Со зголемување на бројот на клетки од пена, се јавува ендотелијално оштетување. Оштетувањето придонесува за активирање на тромбоцитите. Како резултат, тие лачат тромбоксан, кој го стимулира агрегацијата на тромбоцитите, а исто така почнуваат да произведуваат фактор на раст на тромбоцитите, што го стимулира размножувањето на клетките на мазните мускули. Вторите мигрираат од медијалниот до внатрешниот слој на артерискиот wallид, со што придонесуваат за раст на плакетата. Понатаму, зелената плоча со фиброзно ткиво, клетките под влакнеста мембрана се некротични, а холестеролот се депонира во меѓуклеточниот простор. Во последните фази на развој, плакетата е импрегнирана со соли на калциум и станува многу густа. Во областа на плакетата, често се формираат згрутчување на крвта, блокирајќи го луменот на садот, што доведува до акутна циркулаторна инсуфициенција на соодветното место на ткивата и развој на срцев удар. Регулацијата на синтезата на холестерол - неговиот клучен ензим (HMG-CoA редуктаза) се спроведува на различни начини. Фосфорилација / дефосфорилација на HMG редуктаза. Со зголемување на односот на инсулин / глукагон, овој ензим се фосфорилира и преминува во активна состојба. Дејството на инсулин се спроведува преку 2 ензими. HMG-CoA редуктаза киназа фосфатаза, која ја претвора киназата во неактивна дефосфорилизирана состојба: Фосфотаза HMG-Co редуктаза со претворање во дефосфорилизирана активна состојба. Резултатот од овие реакции е формирање на дефосфорилизирана активна форма на редуктаза HMG-CoA. Како резултат на тоа, за време на периодот на апсорпција, холестеролот се зголемува. Во овој период се зголемува достапноста на почетната подлога за синтеза на холестерол - ацетил - CoA (како резултат на јадење храна која содржи јаглехидрати и масти, бидејќи CoA ацетилот се формира при распаѓање на гликоза и масни киселини). Во постсасорбитната состојба, глукагонот преку протеиногеназа А ја стимулира фосфорилацијата на HMG - CoA - редуктаза, претворајќи го во неактивна состојба. Оваа акција е засилена со фактот дека во исто време глукагонот ја стимулира фосфорилацијата и инактивацијата на фосфотазата на HMG-CoA редуктазата, со што ја задржува HMG-CoA редуктазата во фосфорилизирана неактивна состојба. Како резултат, синтезата на холестерол во периодот по апсорпција и за време на постот е инхибирана со ендогена синтеза. Ако содржината на холестерол во храната беше доведена до 2%, тогаш нагло се намали синтезата на ендоген холестерол. Но, целосното престанување на синтезата на холестерол не се јавува. Степенот на инхибиција на биосинтезата на холестерол под влијание на холестерол што доаѓа од храна варира од личност до личност. Ова укажува на индивидуалноста на процесите на формирање на холестерол. Со намалување на интензитетот на синтезата на холестерол, можно е да се намали неговата концентрација во крвта. Ако се расипе рамнотежата помеѓу внесот на холестерол со храна и неговата синтеза во организмот од една страна и излачувањето на жолчните киселини и холестеролот од друга страна, се менува концентрацијата на холестерол во ткивата и крвта. Најсериозните последици се поврзани со зголемување на концентрацијата на холестерол во крвта (хиперхолестеролемија), додека веројатноста за развој на атеросклероза и холелитијаза се зголемува. Семејната хиперхолестеролемија (HCS) - оваа форма е многу почеста - околу 1 пациент на 200 луѓе. Наследниот дефект на HCS е повреда на апсорпцијата на LDL од страна на клетките, а со тоа и намалување на стапката на LDL катаболизам. Како резултат на тоа, се зголемува концентрацијата на ЛДЛ во крвта, како и холестерол, бидејќи има многу од тоа во ЛДЛ. Затоа, со HCS, карактеристично е таложење на холестерол во ткивата, особено во кожата (ксантоми), во theидовите на артериите. Инхибиција на синтезата на HMG-CoA редуктаза Конечен производ на метаболички пат на холестерол. Ја намалува стапката на транскрипција на генот HMG-CoA редуктаза, со што се инхибира сопствената синтеза. Црниот дроб активно синтетизира жолчни киселини од холестерол, и затоа жолчните киселини ја инхибираат активноста на генот HMG-CoA редуктаза. Бидејќи HMG-CoA редуктазата постои по синтеза на околу 3, инхибицијата на синтезата на овој ензим на холестерол е ефикасна регулатива. Погледнете го видеото: Холин есенцијален нутриент за нормална функција на мозокот и телото (Мај 2024).
|